

Research
We investigate the function and regulation of the mTOR signaling pathway to better understand its role in diabetes, aging and cancer.

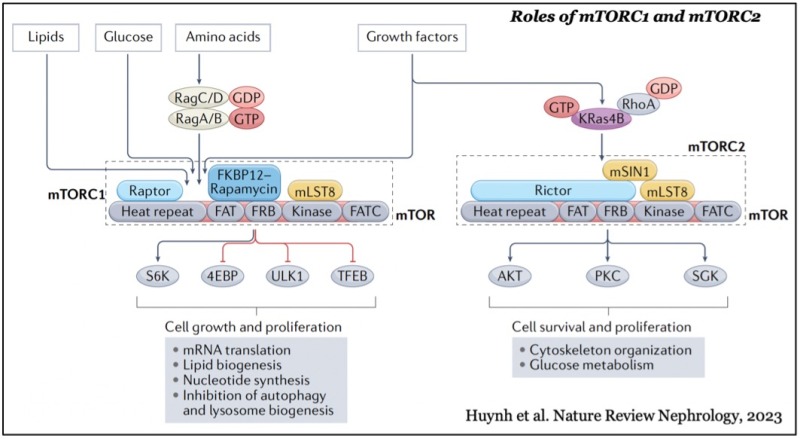
mTOR forms two distinct functional complexes — mTOR complex 1 and 2 (mTORC1 and mTORC2) — which are evolutionarily conserved. mTORC1 exists as a multiprotein complex containing mTOR, Raptor, and mLST8; mTORC2 consists of mTOR, Rictor, mSin1, and mLST8. mTORC1 can be fully activated by both growth factors and nutrients, such as amino acids and glucose on the membrane of lysosomes, a major catabolic organelle that digests unnecessary proteins and damaged organelles. Whereas mTORC1 activity is sensitive to inhibition by rapamycin, an FDA-approved anti-cancer and immunosuppressant, mTORC2 activity is resistant to the drug.
While mTORC1 stimulates major cellular anabolic processes by phosphorylating its dozens of downstream substrates, such as S6 kinase, the reason why its activation occurs on lysosomes, is not fully understood. However, recent studies suggested that mTORC1 may also sense lysosomal luminal amino acids in addition to cytosolic amino acids for its activation, highlighting that the lysosome functions as a platform for both cellular catabolism and anabolism.
In contrast, mTORC2 activity is mainly regulated by growth factors such as insulin and IGF-1 and activated at the plasma membrane. The active mTORC2 phosphorylates and activates AGC-kinases such as Akt and protein kinase C, kinases essential for cell proliferation, survival, cytoskeleton reorganization, and glucose metabolism.
Our studies on the regulation of mTORC1 signaling revealed that the tumor suppressor proteins TSC1 and TSC2 function as negative regulators of mTORC1 (Inoki et al., Nature Cell Biology, 2002). Loss of function of either TSC1 or TSC2 leads to Tuberous Sclerosis Complex (TSC), an inherited tumor syndrome. Tumor formation in the heart, lung, brain and kidney causes severe complications in TSC patients.
TSC1 and TSC2 proteins form a functional complex bearing specific GTPase activating protein (GAP) activity, which inhibits Rheb small GTPase (Inoki et al., Genes & Development, 2003), an essential activator of mTORC1 on the lysosome. Furthermore, we demonstrated that Akt phosphorylates and suppresses TSC2 activity (Inoki et al., Nature Cell Biology, 2002), thereby activating mTORC1 activity. In contrast, another energy-sensing kinase, AMP-activated protein kinase, which stimulates many cellular catabolic processes in response to energy scarce and calorie restriction, also phosphorylates TSC2, resulting in its activation, thereby inhibiting mTORC1 activity (Inoki et al., Cell, 2003).
These studies revealed not only the function of the tumor suppressor TSC proteins but also the mechanism of mTORC1 activation by growth factors. The findings also suggest that rapamycin could be a potential therapeutic drug for TSC patients suffering from multiple tumors. In fact, recent clinical trials confirmed the effectiveness of rapamycin for mitigating tumor development in TSC, and it has been approved to use rapamycin analogs (rapalogs) for the treatment of TSC.
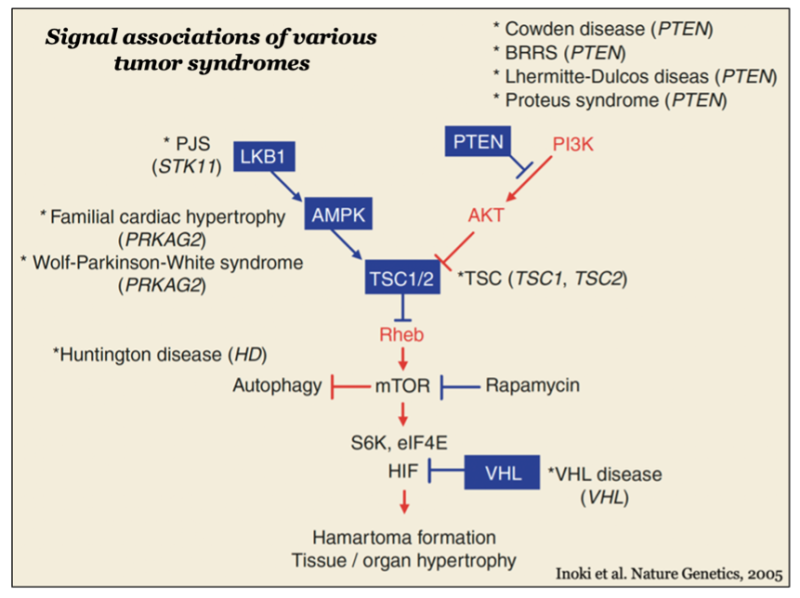
There are various genetic hamartoma and tumor syndromes in which mutations are found in the genes encoding upstream or downstream regulators of the TSC-mTORC1 pathway, such as LKB1 kinase, AMPK, PTEN lipid phosphatase and VHL, suggesting that inactivation of the TSC complex and concomitant mTORC1 activation may be a common molecular etiology in the development of tumors in these syndromes, and rapamycin can be an effective agent for the treatment (Inoki et al., Nature Genetics, 2005).
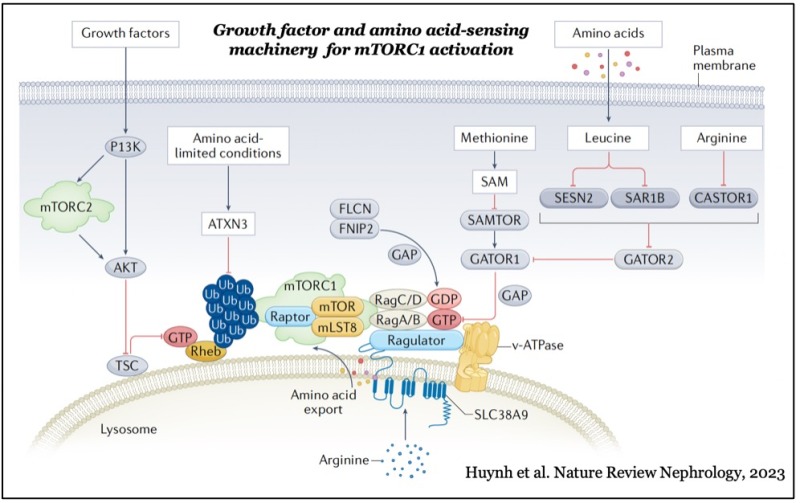
In addition to the growth factor-Akt-TSC-Rheb pathway, the activation of mTORC1 also requires nutrient inputs from amino acids, glucose and lipids. Amino acids (e.g., leucine and arginine) and lipids (e.g., cholesterol and phosphatidic acid) play a key role in activating lysosomal Rag small GTPases, which recruit mTORC1 to the lysosomal membrane where active Rheb induced by growth factors interacts with and stimulates mTORC1 activity.
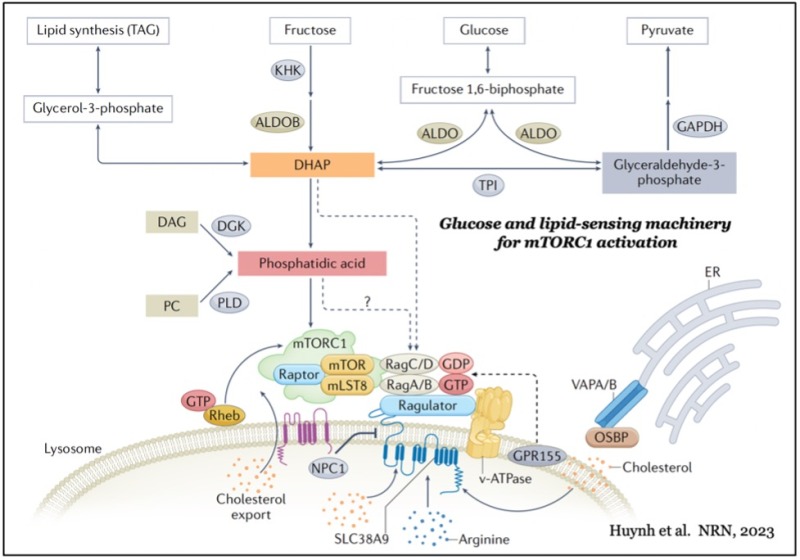
Recent studies identified several key amino acids (e.g., leucine and arginine) and their sensing proteins (e.g., SESN2 and CASTOR1) that regulate Rag small GTPase activity through their cognate GAP and GEF (guanine nucleotide exchange factor) proteins. A particular glucose metabolite (i.e., DHAP), glycolytic enzyme (i.e., aldolase), and lipids such as cholesterol have also been proposed to act as upstream stimulators of Rag small GTPases.
While cholesterol-sensing proteins (e.g., lysosomal GPR155) have been proposed to act as a stimulator for the Rag GTPases, it remains elusive how the glucose metabolite and glycolytic enzyme stimulate lysosomal mTORC1 localization through Rag GTPases. Furthermore, while other phospholipids also enhance lysosomal mTORC1 localization, the molecular mechanisms by which these phospholipids contribute to the recruitment of mTORC1 to the lysosomal membrane have not been identified.
Our laboratory seeks new regulations of nutrient-sensing mechanisms for mTORC1 activation and investigates their roles in metabolic disorders (e.g., obesity, diabetes and its renal complications, non-alcoholic steatohepatitis and cancer) using various cell culture systems and mouse models.
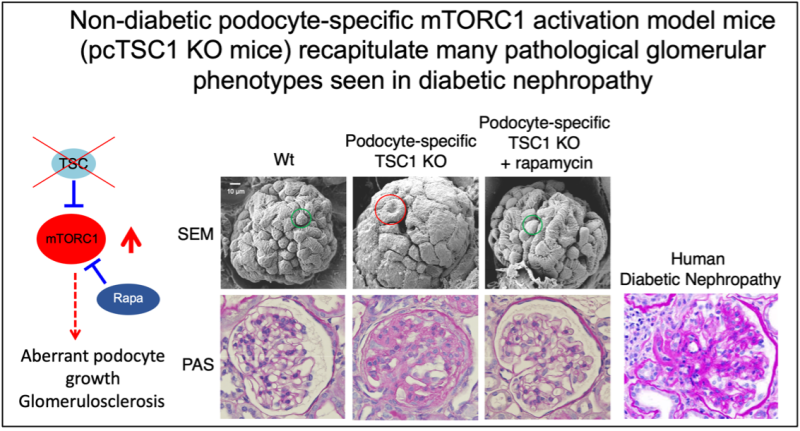
We previously demonstrated that aberrant mTORC1 activation occurs in various tissues — such as the liver, pancreas and kidney — under high-calorie intake and/or diabetic conditions. Importantly, constitutive activation of mTORC1 induced by loss of the functional TSC complex in pancreatic beta cells and renal glomerular podocytes leads to beta-cell exhaustion and podocyte injury, respectively, and mimics their dysfunctions seen in diabetic animal models and patients. Strikingly, the attenuation of hyper-activation of mTORC1 in podocytes of diabetic animals significantly prevented the onset/progression of diabetic kidney disease (diabetic nephropathy: a leading cause of mortality in both type 1 and type 2 diabetic patients), indicating that dysregulated hyperactive mTORC1 in glomerular podocytes contributes to the development of diabetic nephropathy (Inoki et al., Journal of Clinical Investigation, 2011).
However, actual key upstream signals that induce hyperactivation of mTORC1 and crucial downstream of mTORC1 that causes cell dysfunction in diabetes are still not well understood.
Our laboratory investigates molecular etiologies of dysregulated mTORC1 activity in diabetes and obesity using various mouse models and seeks potential therapeutic targets, of which inhibition attenuates and corrects aberrant mTORC1 activation under diabetic and obese conditions.
1. Determine the roles of nutrient-sensing machinery in the development of obesity, diabetes, diabetic complications and NASH.
2. Study the therapeutically targetable pathways that regulate mTORC1 and AMPK in the development of diabetic kidney disease and NASH.
3. Investigate altered organelle-associated components in the specific tissues and cells in various metabolic disorders (e.g., liver and macrophage in NASH, podocyte and tubular cells in obesity and diabetes).