

Research
We study the mechanisms of neural stem cell self-renewal and differentiation, which have implications for neurological disorders and cancer.

Whether through symmetric or asymmetric cell division, stem cell progeny must choose between assuming a stem cell identity by remaining undifferentiated or adopting a progenitor cell identity by committing to differentiate. To successfully adopt a progenitor identity, a stem cell progeny must exit from the stem cell state and stably restrict the developmental potential. In contrast to stem cells, a progenitor cell exclusively generates differentiated cell types. Progenitors with incomplete restriction of developmental potential likely will revert into stem cell-like cells instead, and might be more susceptible to be transformed into tumor-initiating stem cells when acquired tumor driver mutations. Furthermore, ectopically activating tumor stem cells to commit to differentiate can reduce tumor burden. Thus, regulation of developmental potential lies at the intersection of normal programming of stem cell progeny in the healthy state and their abnormal re-programming in the tumorigenic state.
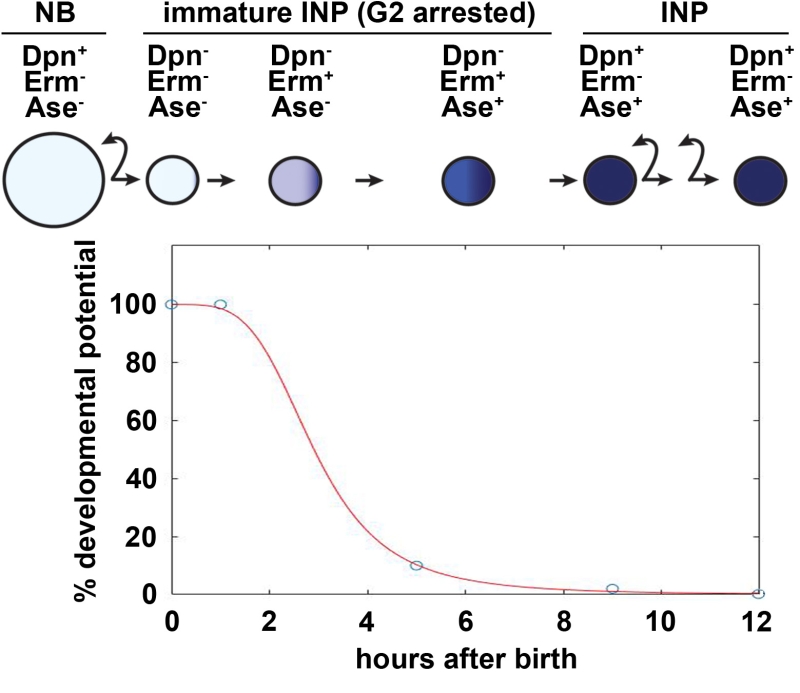
Similar to vertebrate embryonic neural stem cells, Notch signaling plays a key role in regulating the decision to self-renew or to differentiate following asymmetric type II neural stem cell (neuroblast) division in the fly larval brain (Fig. 1). The neuroblast progeny destined to commit to differentiation or the newly born immature INP exits from the neuroblast state and commits to differentiation by initiating the restriction of developmental potential less than 60 minutes after birth. 8 hours later, an immature INP adopts an INP identity and re-enters the cell cycle to exclusively generate differentiated cells. Our lab uses the type II neuroblast lineage as an in vivo paradigm to define the molecular mechanisms that
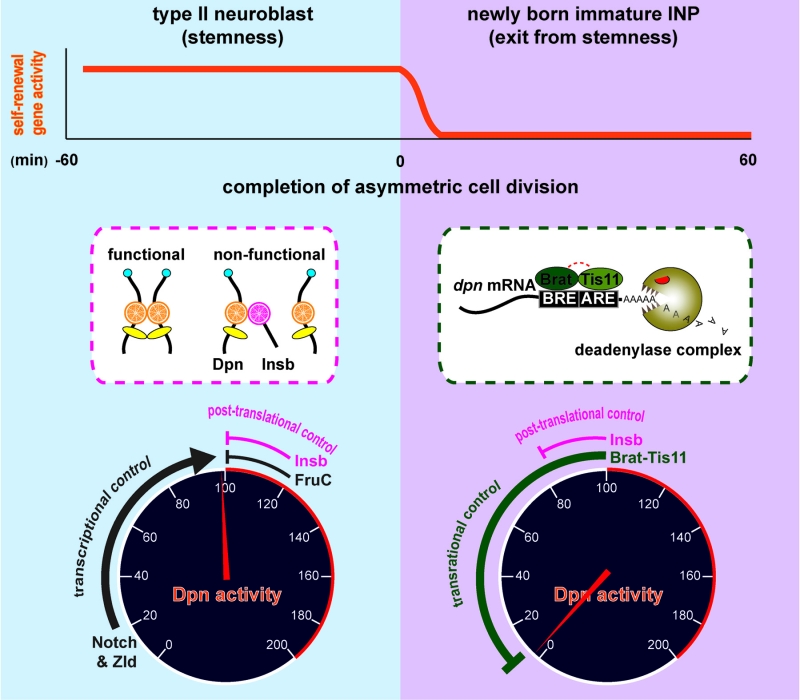
Self-renewal genes maintain stem cells in an undifferentiated state by preventing the commitment to differentiate. Thus, self-renewal gene activity must be down-regulated at all levels to allow stem cell progeny to exit from the stem cell state. In contrast to transcriptional control, little is known about how post-transcriptional regulatory mechanisms terminate self-renewal gene activity. By using the self-renewal gene deadpan (dpn) as a paradigm, we recently demonstrated a multi-layered regulatory mechanism where the synergy between transcriptional and post-transcriptional control terminates self-renewal gene activity in the newly born immature INP. Our proposed multi-layered gene regulation system is likely broadly applicable to the control of exit from the stem cell state in all stem cell lineages and in the regulation of numerous cell fate decisions during normal development.
Our ongoing effort focuses on characterizing (1) how sequence-specific RNA-binding proteins mechanistically link mRNA decay to the termination of self-renewal gene expression, and (2) how multimodal post-translational regulatory mechanisms function through protein degradation and sequestration to terminate self-renewal protein activity.
Rapid transition from exiting the stem cell state to initiating the restriction of developmental potential ensures that stem cell progeny commit to differentiation in a timely manner. Because the master regulators of differentiation trigger the commitment to differentiate, defining the mechanisms that coordinate their activation with the exit from the stem cell state will provide critical insights into the timely initiation of restriction of developmental potential in stem cell progeny. Despite their roles in the commitment to differentiate, the identities of the master regulators remain unknown in most stem cell lineages. As such, the mechanisms controlling the timing of initiation of the restriction of developmental potential remain poorly understood. We previously identified the earmuff (erm) gene that functions as a master regulator of differentiation to restrict the developmental potential of immature INPs. We recently demonstrated that self-renewal transcriptional repressors activated by Notch signaling function through histone deacetylase 1 to maintain erm in a poised state in type II neuroblasts by continually decommissioning its immature INP-specific enhancers. This histone deacetylase-based poising mechanism allows for a rapid onset of Erm-mediated restriction of the developmental potential in the newly born immature INP following the termination of self-renewal factor activity.
Our ongoing efforts focus on (1) investigating the role of chromatin looping in maintaining erm in a poised state in self-renewing neuroblasts, and (2) devising a generalizable model for the histone deacetylase-based poising mechanism.
Stable restriction of developmental potential ensures the generation of a requisite number of differentiated cells during development and homeostasis while reducing the likelihood of the formation of tumor initiating cells during tumorigenesis. Incomplete restriction of developmental potential can allow progenitors cells to spontaneously re-acquire stem cell-like identities, and to serve as the origin of tumor initiating stem cells when acquired tumor driver mutations.
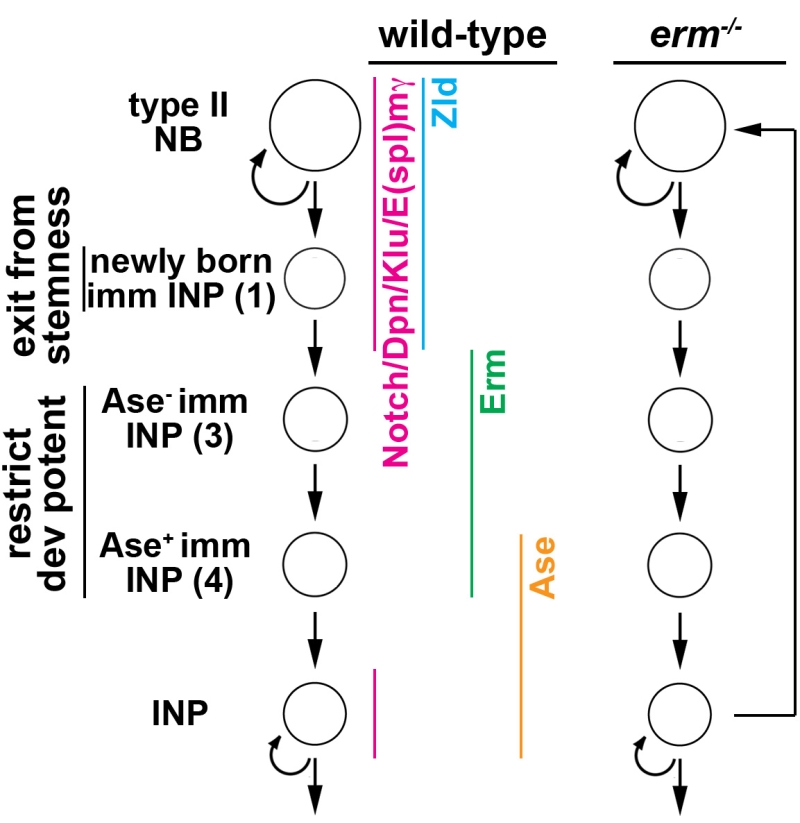
Due to the lack of a well-defined window when the restriction of developmental potential occurs, mechanisms regulating this critical transition remains poorly defined in all stem cell lineages. By using the type II neuroblast lineage as a paradigm, we have identified erm and an additional factor that function together to stably restrict the developmental potential of INPs. Erm likely functions through histone deacetylases to inactivate the transcription of genes required for maintaining the developmental potential. In addition to Erm-mediated histone deacetylation, mechanisms that robustly silence gene transcription must also exist to ensure stable restriction of the developmental potential. Insights into temporal chromatin changes in genes that maintain developmental potential during the commitment to differentiate in immature INPs will likely lead to a generalizable model for stable restriction of the developmental potential in progenitors in all stem cell lineages.
Our ongoing effort focuses on (1) characterizing our newly identified gene that functions together with erm to stably restrict developmental potential, (2) identifying the key downstream target genes critical for stable restriction of developmental potential, and (3) defining the mechanisms by which this gene regulatory system stably restricts developmental potential.